| Adjunto | Tamaño |
|---|---|
| 129.43 KB | |
| 47.39 KB |
{kind=link}
{kind=link}
La última aportación de Víctor Bernal: Hormigas, bosques y territorios, comentada aquí mismo en el blog: http://www.mirmiberica.org/node/339, demuestra que los grupos funcionales, una vez se han simplificado y adaptado al hábitat objeto de estudio son de mucha utilidad y ponen en evidencia la importancia de las hormigas como bioindicadoras.
El estudio teórico del uso de los grupos funcionales ha sido creciente en los últimos años. Los ecólogos, cuando describen la estructura y el funcionamiento de los ecosistemas ponen cada vez más énfasis en el uso de clasificaciones no filogenéticas de organismos (Steffen et al. 1992). Aunque la idea de clasificación funcional se puede remontar a la época de Teofrasto (372-287 a. C.), el debate moderno comenzó a partir de la década de los 60 del siglo pasado (Gitay & Noble, 1997). Estos autores, junto con Terborghi & Robinson (1986), Hawkins & MacMahon (1989) y Simberloff & Dayan (1991), revisan las diferentes definiciones y la creciente literatura sobre clasificaciones gremiales, grupos de respuesta y grupos funcionales, y su amplio abanico de aplicación la ecología.
Para conocer mejor la organización de las comunidades animales, podemos dividir sus niveles tròficos en "gremios" (guilds). Históricamente, fue Root (1967) quien introdujo el concepto ecológico de los gremios, que definió como "un grupo de especies que explotan por igual los mismos recursos ambientales o bien las especies que se solapan significativamente en los requerimientos de su nicho". Es decir, los gremios son grupos de especies que explotan un recurso básico común de una manera parecida. Posteriormente, muchos autores han propuesto diferentes definiciones y agrupaciones. Terborghi & Robinson (1986) considera que los grupos funcionales también consisten en gremios específicos de especies que explotan los mismos recursos, pues en la práctica, estos gremios casi siempre se constituyen en función de las relaciones tróficas como por ejemplo las aves insectívoras (Root 1967), las especies granívoras del desierto (Brown JH & Davidson 1977) o los insectos herbívoros (Moran & Southwood 1982).
Pero en el caso de las hormigas, donde la mayoría de especies tienen requerimientos similares de recursos, el concepto trófico de los gremios no es demasiado útil (Andersen 1991). Por tanto, las agrupaciones funcionales propuestas para las plantas, basadas en un amplio abanico de caracteres ecológicos, son más apropiadas para la agrupación funcional de las hormigas, que no las de los animales (Andersen 1995). Progresivamente se ha ido diferenciando el concepto de gremio del de "grupo funcional": según Gitay & Noble (1997) un grupo funcional, que podemos considerar sinónimo de tipo funcional, se define y se diferencia del resto de conceptos mencionados, y agrupa a los organismos que responden a una perturbación de la misma manera y mediante el mismo mecanismo (Tabla).

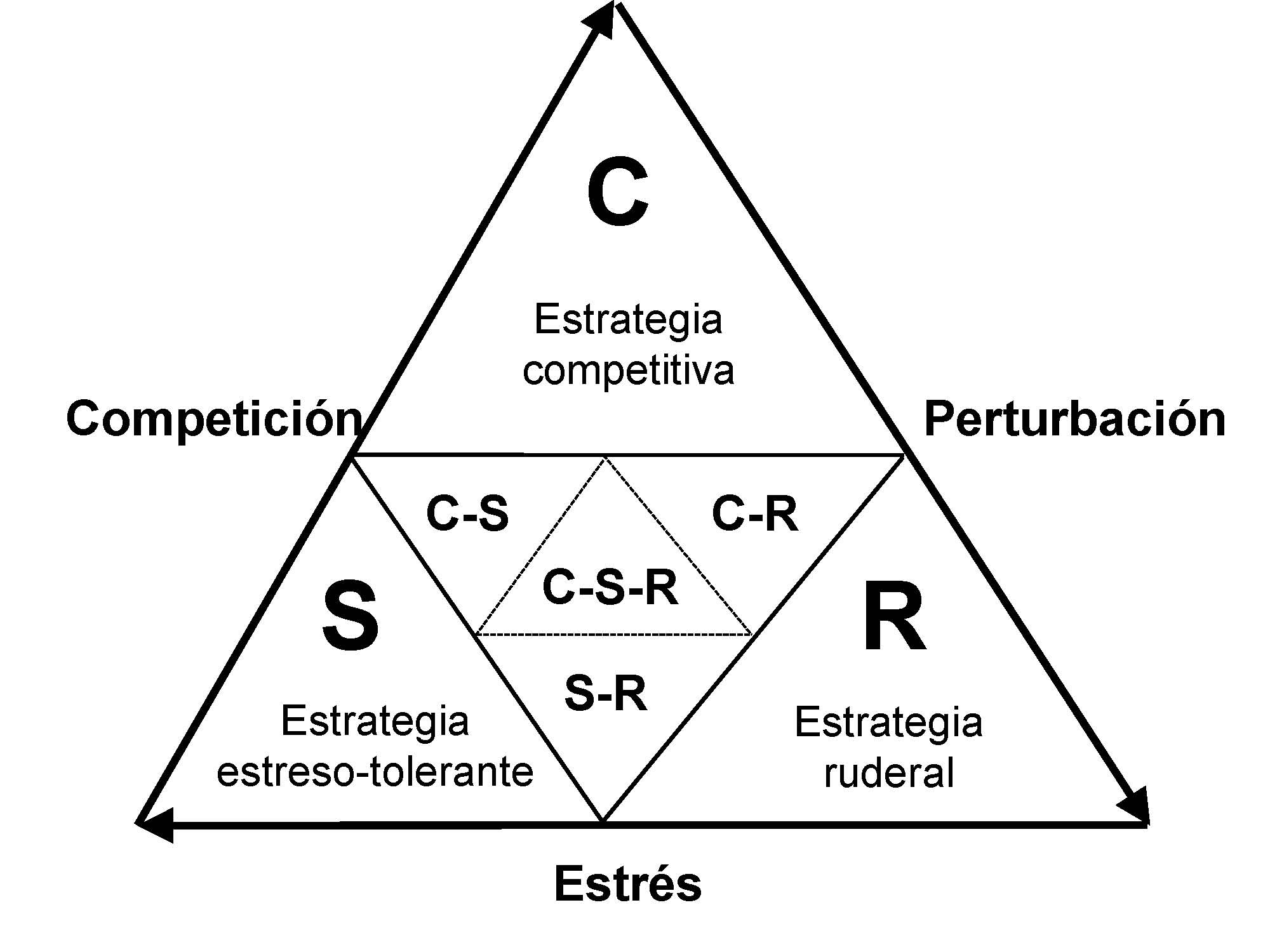
Inicialmente, el concepto de grupo funcional se desarrolló como una continuación del que se había aplicado a las comunidades vegetales (Grime 1974, 1979; Figura), donde desde una perspectiva global se clasifican de acuerdo con la importancia relativa los factores más determinantes de la vegetación (Lavorel et al. 1997; Smith et al. 1997): el estrés (cualquier factor limitante de la productividad) y la perturbación (cualquier factor que disminuya la biomasa). Así, con la intención de simplificar la aparente complejidad de los sistemas ecológicos y, basándose en criterios ecológicos, los investigadores han agrupado varias especies en unos grupos funcionales, que trascendiendo los límites taxonómicos y biogeográficos, responden de manera predictiva al estrés y a la perturbación (Andersen 2000).
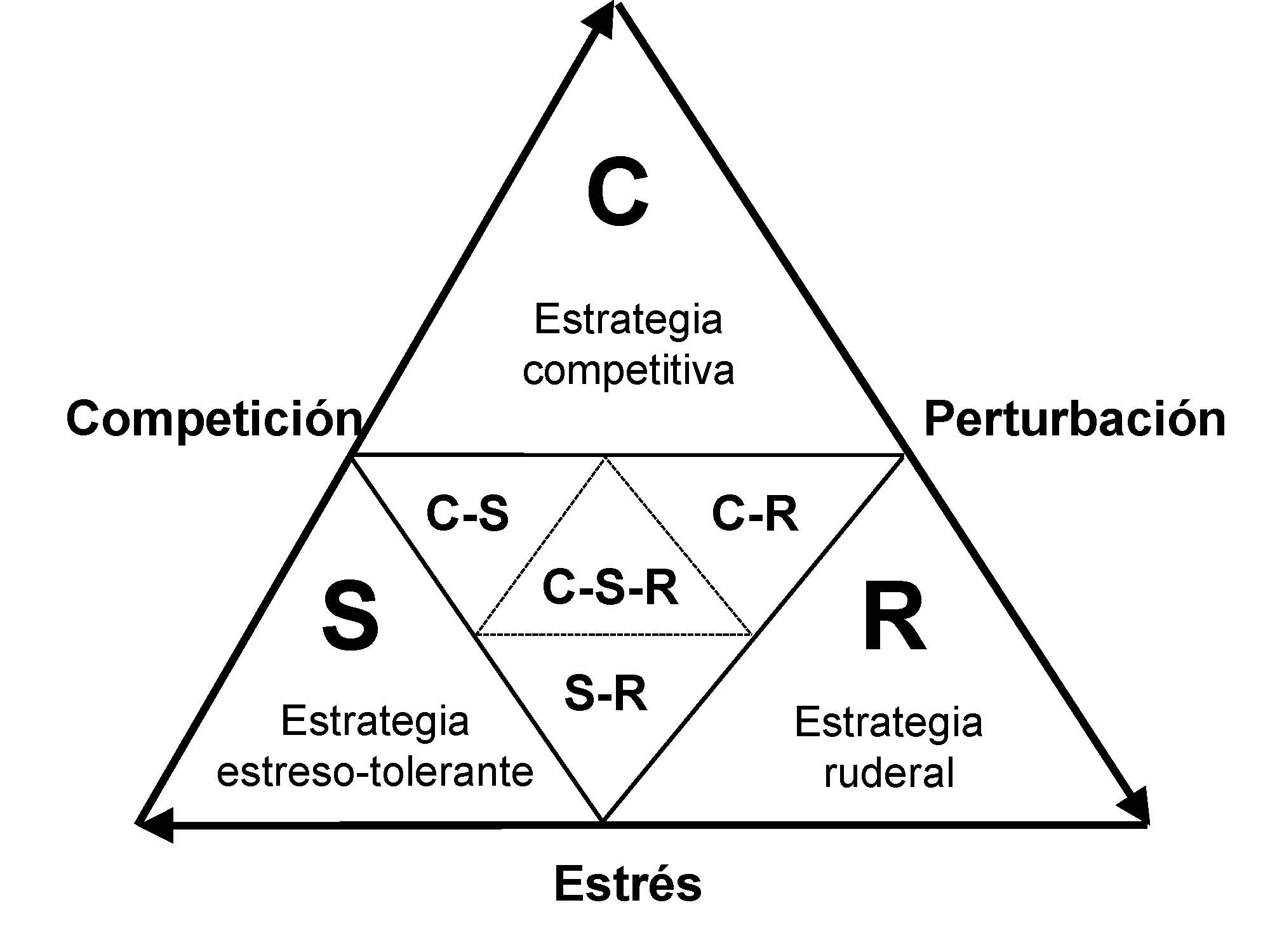 Figura. Clasificación de las comunidades vegetales en relación al estrés y la perturbación, siguiendo la nomenclatura de Grime (1974, 1979). Modificado a partir de Andersen (2000)
Figura. Clasificación de las comunidades vegetales en relación al estrés y la perturbación, siguiendo la nomenclatura de Grime (1974, 1979). Modificado a partir de Andersen (2000)
En los estudios que utilizan las hormigas como organismos bioindicadores, cada vez se usa más la agrupación de los taxones en unos grupos funcionales que engloban las especies mediante criterios que tienen en cuenta más el comportamiento ecológico que no la afinidad taxonómica, convirtiéndose , por tanto, una herramienta muy potente para avanzar en la comprensión de los sistemas ecológicos, ya que facilitan un análisis más cuidadoso y práctico de los cambios observados (Andersen 1990, 1995). Como que la abundancia de los organismos integrantes de los grupos funcionales está estrechamente ligada al estrés y a la perturbación, su utilización permite situar las diferentes especies de hormigas en su contexto ecológico específico, simplificando las aparentemente complejas pautas de distribución de especies, y mejorando, las medidas tradicionales de biodiversidad, como la riqueza, diversidad y abundancia (Andersen 1997). Según el nivel de perturbación, unos grupos funcionales pueden ser dominantes y otros, los que se adapten peor, disminuir o desaparecer (Stephens & Wagner 2006).
Aspectos conflictivos a debatir y analizar:
Desde mi punto de vista, y aquí empieza tal vez el primero de los debates, la aplicación de los grupos funcionales a las hormigas presenta una dificultad inicial e histórica sobre si estos grupos funcionales deben (pueden) aplicarse homogéneamente a todas las zonas biogeográficas o bien debería haber una batería de grupos funcionales específica para cada región o hábitat. Digo histórica pues hasta el momento ha sido unánime la aplicación (en algunos casos la adaptación parcial) de los grupos funcionales definidos inicialmente para todas las áreas de estudio ya fueran éstas de Australia y de Norteamérica o bien del área Mediterránea. Con ello, desde luego, se consigue una comparabilidad y compatibilidad global entre los diferentes estudios pero, a mi parecer, a base de forzar en demasía los integrantes del grupo funcional en cuestión.
El segundo aspecto a analizar es si el original y principal punto de vista comportamental (básicamente la dominancia - subordinación) adoptado para los primeros grupos funcionales definidos en Australia son aplicables al área Mediterrànea, zona biogeográfica de nuestro interés principal.
En mis próximas aportaciones intentaré presentar los pros y los contras y mi punto de vista acerca de ambos aspectos.
Referencias
Andersen AN (1990) The use of ant communities to evaluate change in Australian terrestrial ecosystems: a review and a recipe. Proceedings of the Ecological Society of Australia 16:347-357
Andersen AN (1991) Parallels between ants and plants: Implications for community ecology. In: Huxley CR, Cutler DC (eds) Ant-Plant interactions. Oxford University Press, Oxford, pp 539-558
Andersen AN (1995) A classification of australian ant communities, based on functional-groups which parallel plant life-forms in relation to stress and disturbance. Journal of Biogeography 22: 15-29
Andersen AN (1997) Using ants as bioindicators: Multiscale issues in ant community ecology. Conservation Ecology (on-line) 1:8
Andersen AN (2000) A Global Ecology of Rainforest Ants: Functional Groups in Relation to Environmental Stress and Disturbance. Chapter 3. In: Agosti D, Majer JD, Alonso L, Schultz R. (eds) Standard Methods for Measuring and Monitoring Biodiversity. Smithsonian Institution Press, Washington, pp 25-34
Brown JH, Davidson DW (1977) Competition between seed-eating rodents and ants in desert ecosystems. Science 196:880-882
Gitay H, Noble IR (1997) What are functional types and how should we seek them? In: Smith TM, Shugart HH, Woodward FI (eds) Plant functional types their relevance to ecosystem properties and global change. Cambridge University Press, Cambridge, pp 3-19
Gitay H, Noble IR (1997) What are functional types and how should we seek them? In: Smith TM, Shugart HH, Woodward FI (eds) Plant functional types their relevance to ecosystem properties and global change. Cambridge University Press, Cambridge, pp 3-19
Grime JP (1974) Vegetation classification by reference to strategies. Nature 250:26-31
Grime JP (1979) Plant Strategies and Vegetation Processes. John Wiley and Sons, Chichester, UK
Hawkins EM & MacMahon JA (1989) Guilds: multiple meanings of a concept. Annual Review of Entomology 34:423-451
Lavorel S, McIntyre S, Landsberg J, Forbes TDA (1997) Plant functional classifications: from general groups to specific groups based on response to disturbance. TREE 12:474-478
Moran VC, Southwood TRE (1982) The guild composition of arthropod communities in arid Australia and North America. Ecological Monographs 58:19-38
Root RB (1967) The Niche Exploitation Pattern of the Blue-Gray Gnatcatcher. Ecological Monographs 37:317-350
Simberloff D, Dayan T (1991) The guild concept and the structure of ecological communities. Annual Review of Ecology and Systematics 22:115-143
Smith TM, Shugart HH, Woodward FI, eds. (1997) Plant functional types their relevance to ecosystem properties and global change. Cambridge University Press, Cambridge
Steffen WL, Walker BH, Ingram JSI, Koch GW eds. (1992) Global Change and Terrestrial Ecosystems: the Operational Plan. IGBP-ICSU, Estocolm
Terborgh J, Robinson S (1986) Guilds and their utility in ecology. In: Kikkawa J, Anderson DJ eds. Community ecology pattern and process. Blackwell Scientific Publications, Melbourne. pp. 65-90